August, 2017
Page after page, we have been pondering patterns and processes in community ecology under the sounds of gusting winds and heavy rain. From one storm to the next, when our field days were cut short, we could sit by the fire in the Community Building (the oldest building in the Yukon) and delve in deeper into Mark Vellend’s “The Theory of Ecological Communities”.

We talked about selection and the many selection-focused hypotheses of shaping and maintaining ecological communities, and singled out the ones most relevant to the tundra and our work. We then considered speciation, dispersal and drift, which still influence communities on the island, though they might not be as important relative to selection.
We have now wrapped up our field season and returned to Vancouver. Along with mailing away samples and inventorising equipment, we have also turned the final pages of Mark’s book. Here, we will share our final book club thoughts, with a particular focus on the theories, which relate most to our work in the tundra.




We see our study of vegetation change in the Arctic as being detective work – you figure out a question, gather evidence, open the data present, and interpret what it means. For us, explanation and prediction in ecology go hand in hand, and we aim to do both – you can’t make predictions without knowing how your system works. Although theory development is perhaps not the main motivation of our work, we don’t shy away from reading about different theories and seeing how they can inform our research. In fact, we go as far as playing “Guess the ecological theory” at breakfast – a stimulating start to the day!
We think of theory as a way to simplify complex ecological phenomena in order to make them generally applicable. The same rules don’t apply everywhere, though, and our goal is to find which rules are relevant for the tundra biome, not necessarily to extrapolate across the entire world. The same global change drivers have different impacts across the world’s biomes, making the drawing of universal conclusions even harder. Nevertheless, sometimes theory works, and here, we will present the Arctic edition of when that happens.
Table 1. Theories we consider relevant to tundra vegetation change and our work.
| Theory | Links to the tundra biome and our work | |
| The competitive exclusion principle (Gause 1934) and R* theory (Tilman 1982) | Competitive exclusion likely exists within the tundra biome with species competing for the limited resources of the tundra environment. Species have specific adaptations from forming tussocks to outcompete other species and form their own microclimate to having evergreen leaves allowing them to start photosynthesis as soon as the snow melts in spring. Each plant species has its own combination of traits to deal with the short growing seasons of the Arctic and those different traits likely promote coexistence. Each species likely does have its single resource for which it can survive at the lowest equilibrium level (R*), but figuring out which species are best adapted to exploiting which resource is a bit of a challenge. | |
| Temporal and spatial storage effects (Chesson 2000b) – Selection spatially variable and negatively frequency dependent | The spatial and temporal storage effect is likely at play in a major way the Arctic. Species are specifically adapted to the harsh environmental conditions and can store resources for reproduction or growth during hard times that they can then use when conditions allow. Some species are adapted to thriving under snow patches, others to coping with drought conditions, others to dealing with flooding and soil saturation yet all can cope with the cold. These different sorts of adaptation can promote species coexistence and diversity across the tundra landscape. | |
| Enemy-mediated coexistence (Holt et al. 1994) | Enemies do exist in the tundra. We have been monitoring herbivory sign for the past four years on the island and have found the rates of herbivory to be relatively low. But the herbivores are out there and are a selective pressure in the landscape. From leaf herbivores from caterpillars to lemmings to caribou, seed and fruit herbivores from fly larvae to humans, herbivores can take a bite out of the carefully allocated resources that tundra plants have invested in growth and reproduction. Likely herbivory is a bigger deal in places like the European Arctic or Northern Quebec where large herds of reindeer or caribou roam around munching on shrubs. The main Porcupine Caribou heard doesn’t make it out to Qikiqtaruk and large animal herbivory rates are lower. How important is herbivory at sites across the Arctic? Well that is something that the Herbivory Network is trying to figure out. | |
| Priority effects (reviewed in Fukami 2010) | Do initial colonists get an advantage in the tundra? Perhaps this is particularly relevant in the Herschel vegetation type where disturbance rates are lower and the species that invade first might be able to persist longer. | |
| Multiple stable equilibria (reviewed in Scheffer 2009) | Are there multiple stable states in the tundra? This theory is probably quite relevant for the transition from sedge- or herb- to shrub-dominated tundra as tundra shrubification occurs as can be seen in repeat photographs on Qikiqtaruk. | |
| Succession theory (Pickett et al. 1987) | Succession is at play across the tundra biome where disturbance is likely the major factor shaping ecological communities. | |
| Intermediate disturbance theory (Grime 1973, Connell 1978) | The intermediate disturbance theory can be tested with our data – check out our plot of bare ground (a proxy for cryoturbation, a type of disturbance ) vs species richness at the plot level in 2017 (Figure 1). Depending on how you look at it we either do or don’t see the expected hump-shaped relationship of species richness in relation to disturbance – in our case cryoturbation. | |
| Metacommunities: species sorting (Leibold et al. 2004) – spatially variable selection, different species are at an advantage under different environmental conditions | Species sorting and spatially variable selection due to different environmental conditions might explain the different plant communities that we see on the island. See the Book Club II blog post for more thoughts on selection. | |
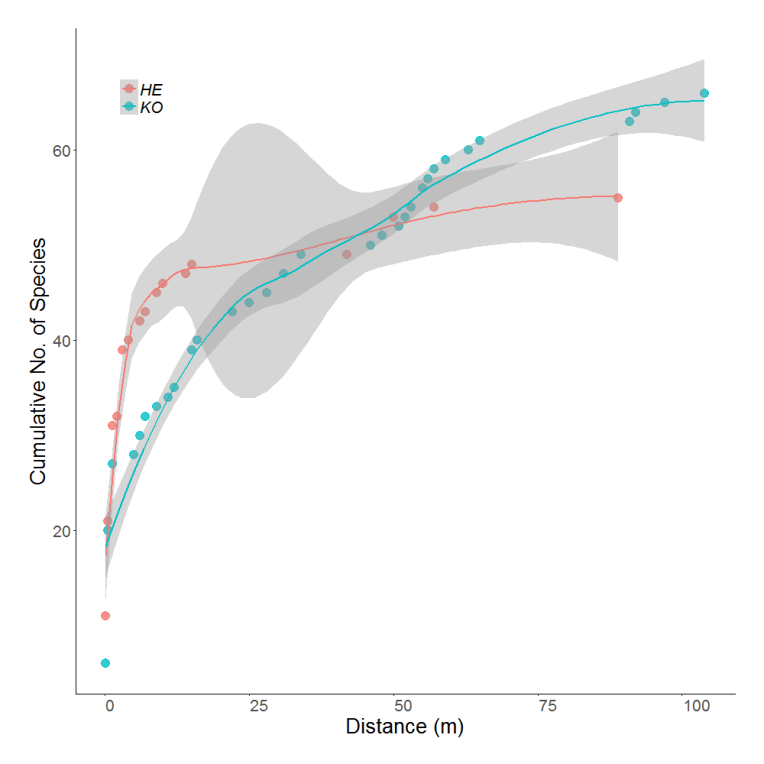
| The species pool hypothesis (Taylor et al. 1990) | Different species pools sizes on the island or at a larger spatial scale within the tundra biome likely do influence local richness. Through the ArcFunc project we can test this theory further. You can also check out our species-accumulation curves (Figure 2). | |
| Island biogeography (MacArthur and Wilson 1967) | Island Biogeography can be tested using breakfast cereal, so it probably is at play in the tundra biome with distance from glacial refugia being a key factor. Isolation could explain the species richness and ecological communities on Qikiqtaruk relative to the adjacent mainland. And certainly, island isolation creates barriers limiting the expansion of tall shrub species within the Arctic at the biome scale. | |

Our work on tundra vegetation change spans across several different levels of analysis, which together will hopefully shed light on how communities in the Arctic are responding to global change drivers. We are monitoring particular individuals for phenology, and surveying 12 x 1 m2 community composition plots on one Arctic island, four pairs of 100 m2 plots for our drone surveys (in the Herschel and Komakuk communities), large-extent sites part of the HiLDEN network, and finally the tundra biome, which we can study as a whole thanks to the ITEX and ShrubHub networks. We are investigating how community composition is changing by studying species and traits in collaboration with the Tundra Trait Team. Along the way, we often come across what Mark calls the “three-box problem” – there will probably always be silent or ‘lurking’ variables we fail to account for, and drawing the line between causation and correlation is rarely easy.
Do we observe different species in a particular place because the environment is different, or is the environment different, because the species are different? What is the trade-off between diversity and invasibility? This year, we recorded the same number of species (32) in both the Herschel and Komakuk plots, but the Komakuk plots are probably more susceptible to invasions, since with more bare ground patches, there is more room for new species to germinate and establish. Alopecurus alpinus has already taken advantage of this opportunity, and more species might follow soon, as from our species pool survey, we found that there are many more species found close by to our plots in both the Komakuk and Herschel vegetation types.







Having read “The Theory of Ecological communities”, spent many hours out in the field and conducted preliminary analyses of our collected data, we think that species sorting and the metacommunity framework, applies best to Qikiqtaruk-Herschel Island. Here, we can observe very strong spatially variable selection, leading to the establishment and presence of distinct vegetation communities. Although of relatively less importance, patch dynamics could also be at play – spatial and temporal variation in selection could cause local extinction, and dispersal could lead to (re)colonisation of certain species. Such is the case with the grass species Alopecurus alpinus that colonised our Komakuk community composition plots between 2004 and 2009. The neutral framework, on the other hand, has little application in the Arctic context, because drift is not a dominant force shaping ecological communities here. Mass effects, whereby selection is spatially variable and not strong enough to prevent immigrants from establishing sink populations, is also likely of little relevance to the tundra, since selection here is very strong, potentially preventing new immigrants from establishing in the first place.

We thoroughly enjoyed reading “The Theory of Ecological Communities” whilst on fieldwork at our remote field site in the Canadian Arctic. There is particular charm in reading about a certain ecological process, be it high- or low-level, and then observing it in action moments later in the field. We look forward to continued discussions of the synthesis of ecological theory, but definitely agree with Mark that four high-level processes do shape community composition – selection, speciation, dispersal and drift.
By Gergana and Isla
_____________________________________
This blog post was written on Qikiqtaruk-Herschel Island in the Western Canadian Arctic as part of Team Shrub’s island book club, aiming to read and discuss Mark Vellend’s 2016 book “The Theory of Ecological Communities” while we are out in the field, right next to the communities we study. Team Shrub are a group of plant ecologists who often work in high-latitude tundra ecosystems on topics in community ecology.
The team’s book club discussions are summarised in four blog posts:
- Qikiqtaruk Book Club Part I: Ecological communities in the Arctic
- Qikiqtaruk Book Club Part II: Selection in the Arctic
- Qikiqtaruk Book Club Part III: Speciation, drift and dispersal in the Arctic
- Qikiqtaruk Book Club Part IV: Theory and high-level processes in the Arctic

8 thoughts on “Qikiqtaruk Book Club Part IV: Theory and high-level processes in the Arctic”